- 果樹栽培歴5年目になる僕が徒然なるままに書くコラム.
- youtubeのチャンネル登録者5.28万人「農の共有 ~けんゆーの農ライフ!~」.
- 色々な相談が来るので,このブログで回答していくよ.

こんにちは.けんゆー(@kenyu0501_)です.
感想ツイートや引用ツイートなど歓迎!
「The Avocado botany production and uses」は,アボカド栽培をする人らが絶対に数回は読んでおく神書だと感じるが,英語であることと,文字量が膨大であることで読むのにまとまった時間が必要だと思われる.そのため,移動時間などに手軽にスマホで読めるととても嬉しい.
本書は,第1章〜第15章で構成されるが,各章をそれぞれ,日本語訳にしたものをこちらで紹介したい.
まずは第1章「イントロダクション」である.
著者は, B. Schaffer、B.N. Wolstenholme、A.W. Whiley.
ちなみに,こちらは自分用のメモ.
普段使っているChatGPTとの会話のメモのうち,価値があると思ったものを残している.
日本語訳は大体はChatGPTがやってくれた.読まれる方は,勝手に参考にしてね.
イントロダクション
アボカド (Persea americana Mill.) は、Laurales 目の Lauraceae 科に属する植物であり、この科は約 50 属、2500 から 3000 種の主に樹木と一部の低木を含む大規模な熱帯植物のファミリーです (Rowher, 1993)。このファミリーには、経済的および文化的に重要な種が含まれ、例えば月桂樹 (Laurus nobilis L.) があります。古代ギリシャでは、月桂樹の葉が功績を称えるために捧げられ、現代でも「ローレート」という言葉が使われています。シナモン (Cinnamomum) の属には、商業的に重要なシナモンとカンフルの木が含まれ、Aniba からは香料油が得られます。Chlorocardium、Eusideroxylon、Persea、および Beilschmiedia の属には重要な木材の木が含まれます。Chanderbali ら (2008) によれば、アボカドは Lauraceae 科の中で最も重要な商品です。世界のアボカド果実の生産量は現在 350 万トンを超えており、そのうち約 20% が国際貿易されています。
進化の側面 – ‘Evo-devo’ と ‘Evo-eco’
Lauraceae 科は、長い間「原始的」または比較的特殊化していない被子植物のファミリーと見なされてきました。この科の花は、多くの小さな、区別のない萼片と花弁を持ち、劣性の子房を持っています。現代の分類学的手法を使用して、Chanderbali ら (2008) は、P. americana (Laurales) が、Soltis および Soltis (1990) によって記述された広く受け入れられている被子植物の系統樹の基部付近に位置する、Magnoliids の系統的に重要な位置にあることを指摘しています。このクレードには、古い分類の「原始的」または「基部」被子植物のほとんどが含まれます。実際、アボカドは、非花植物 (裸子植物) から被子植物の花の進化を理解するための遺伝モデル植物として確立されています (Chanderbali et al., 2008, 2009)。
世界のアボカドコミュニティは、主要な分類学者、進化生物学者、植物学者、および園芸学者がアボカドを最先端のゲノム研究のモデル種として見なしていることに恵まれています。トランスクリプトームシーケンス、ゲノムマッピング、および部分的なゲノムシーケンスなどの現代のゲノム研究技術は、アボカド全ゲノムのシーケンスへの大きな一歩となります (Chanderbali et al., 2008)。生産者にとっての潜在的な利益には、改良された栽培品種と台木、品質の高い果実のより規則的な収量の向上、および主要な病害虫に対する耐性の向上 (特に Phytophthora 根腐れ病) が含まれます。
Lauraceae 科は、Laurales の現存する種の大部分を提供し、中生代白亜紀中期に特に広範囲かつ豊富でした。彼らは高い基本染色体数を持ち (アボカドの 2n = 24)。すべての現存する Lauraceae は、少なくとも 1 億年前の古代の倍数性イベント (s) に起因すると信じられており、Lauraceae での 2 回目の後期倍数性の発生がありました。アボカドの花の進化と花の発達研究 (‘evo-devo’) は、本書第 3 章の Chanderbali らによって要約されています。彼らは、アボカドが花の進化に関するさらなる研究の魅力的なモデルであると結論づけています。アボカドの果実は、ボタニカルな「ベリー」であり、いくつかの独特の特徴を持っています。これには、樹上保存 (前クライマクテリック) の可能性が含まれます。果実がまだしっかりと木に付着している間の継続的な細胞分裂。熟成と共に顕著な呼吸クライマクテリック。したがって、この果実は、果実の熟成の上流規制に関する遺伝子プロファイリング研究にも適しています。
大型のメソアメリカン森林果実の進化生態学 (‘evo-eco’) に関する画期的な論文 (Janzen and Martin, 1982) は、Wolstenholme と Whiley (1999) のアボカド進化生態学に関する推測的な園芸的議論のインスピレーションとなりました。このテーマは、Barlow (2000) の書籍『The Ghosts of Evolution』で詳述されています。人類がアメリカ大陸に到達したのは約 13,000 年前ですが、この栄養価の高い油貯蔵果実は、熱帯低地および熱帯高地のメソアメリカの森林で長い進化の歴史を持っています。Janzen と Martin (1982) は、今では絶滅した更新世の大型動物が特定の森林果実に引き寄せられ、全体を飲み込み、種子にほとんど損傷を与えることなく効果的に分散した可能性が高いことを指摘しました。これには、象に似たゴンフォテリウム、トキソドン、巨大地上ナマケモノ、およびグリプトドンが含まれます。彼らは、数百万年にわたって高エネルギーと栄養ニーズを持つ動物と共進化してきました。本書第 2 章では、Bost らがアボカドの歴史に関する詳細な説明を提供しています。
Barlow (2000) は、アボカドが商業的に栽培されている果実の中で、世界で最も栄養価の高い果実であると結論付けました (Purseglove, 1968)。過去約 11,000 年間 (大型草食動物が絶滅して以来)、それは進化の「幽霊」であり、人間による分散にほぼ完全に依存していました。この独特の果実は、メソアメリカのその生息地およびその近くで少なくとも 9,000 年間、先住民によって評価され、利用されてきました (Smith, 1966; Gama-Campillo and Gomez-Pompa, 1992; Chanderbali et al., 2008)。
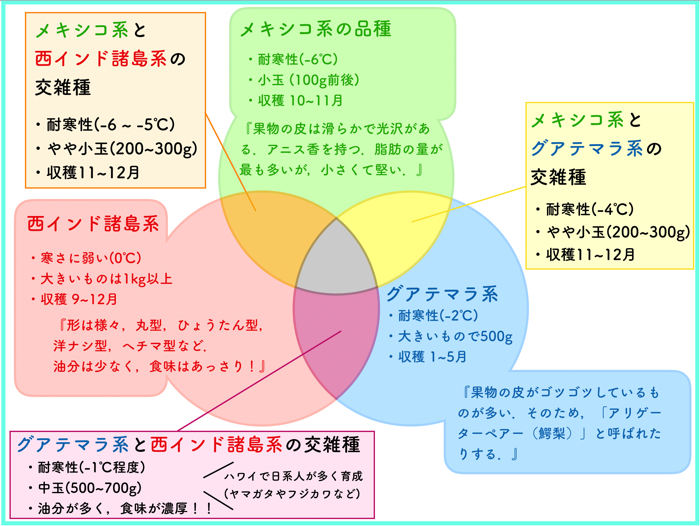
アボカドは、マヤ文明およびアステカ文明によっても評価されており、そのアイコノグラフィー (絵文字) によって証明されています (Storey et al., 1986; Gama-Campillo and Gomez-Pompa, 1992)。これらの文化が、果実の大きさを選択し、食用の質を向上させ、推定される自然範囲外の新しい地域に果実を徐々に広めたと信じられています。数千年にわたる遺伝子の混交と半家畜化が行われました。今日、非常に変異に富んだアボカドは、熱帯低地の西インド種 (誤称)、熱帯高地のメキシコ種 (または「冷温帯」) および熱帯高地のグアテマラ種 (または「温暖温帯」) に園芸的に分類されます。優れた栽培品種の選択と栄養繁殖は、110
年前の米国フロリダでの「ポロック」および「トラップ」に始まりました。ほとんどの現代の栄養繁殖された「温帯」栽培品種は、メキシコ種とグアテマラ種の部分的な雑種です。「フエルテ」は、1911 年に苗木の台木に接ぎ木された最初の「優秀な基準」でした。その後、数百の他の品種が選ばれ、リリースされました。今日の温帯の基準である「ハス」は、1950 年代以来広く栽培されています。フロリダでも、熱帯品種は主に 100 年以上にわたって選ばれ、栄養繁殖されています (本書第 8 章の Crane らおよび第 9 章の Ernst らを参照)。商業農園の要求に関しては、アボカドはまだ完全に家畜化されるまでには遠い道のりがあります。この点で、それは比較的新しい果実です。
アボカドの世界的な果実としての地位
Samson (1986) は、年間生産量に基づいて果樹作物を 4 つのグループに分類しました:
1. 年間生産量が 1,000 万トンを超えるもの (例: ブドウ、柑橘類、バナナ、リンゴ、プランテン、マンゴー)。
2. 年間生産量が 100 万トンから 1,000 万トンの間のもの (例: ナシ、アボカド、パパイヤ、モモ、プラム、パイナップル、デーツ、イチジク)。
3. 年間生産量が 10 万トンから 100 万トンの間のもの (例: カシューナッツ)。
4. 信頼できる統計がない果実 (例: グアバ、ブラジルナッツ、ライチ、マカダミア、サワーソップ)。
1950 年以来、ブドウ、ナシ、およびモモの生産量は倍増し、柑橘類とバナナは 3 倍、パイナップルは 6 倍に増加しました。アボカド、キウイフルーツ、およびライチは「急速な発展」を経験しました。2011 年の年間生産量 (単位: 百万トン) は、柑橘類が約 125、バナナが約 93、リンゴが約 70、マンゴーが 35 以上、ナシおよびモモがそれぞれ約 20、アボカドが 3.5 以上でした (Naamani, 2011)。したがって、世界の生産量の観点から、アボカドはまだ第二段階の果実であり、一般的には主要な「でんぷん質の主食」としては消費されないため、今後もその地位を維持する可能性が高いです。それでも、この果実がヨーロッパのほとんどの地域や急速に工業化しているアジアの一部でよりよく知られるようになるにつれて、比較的低い基盤からの大きな成長の余地があります。さらに、アボカドの食事価値 (主に一価不飽和脂肪酸 (油) を含む) と、例外的なミネラル、ビタミン、および他の有益な抗酸化物質の含有量は、ますます認識されるようになっています (本書第 2 章の Bost らを参照)。
近代的な農園への適応
生態学的側面
アボカドのメソアメリカ起源には、熱帯低地タイプ (いわゆる西インドまたはアンティル種) アボカドの場合、海抜ゼロから数百メートルの高度まで、および高地メキシコおよびグアテマラ (「亜熱帯」) 種アボカドの場合、最大 3000 メートルまでの生息地が含まれます。原生生息地の山岳地帯の特性により、気候および土壌タイプの幅広い範囲が生じ、大部分は夏にピークを迎える高降雨地域にあります。したがって、アボカドの遺伝的多様性が驚くべきことではなく、過去の気候変動に対応して山脈を上り下りする進化的移動があったに違いありません。
P. americana は、主に地理的に熱帯緯度 (北緯 23.5 度から南緯 23.5 度) 内で進化したと考えられており、したがってアボカドは「熱帯果実」としてリストされることがよくあります。しかし、これは純粋に地理的なものであり、誤解を招く可能性があります。Storey ら (1986) は、原産地を北緯約 24 度から北緯 8 度までとしていました。園芸的および気候的には、西インド種のみが低地の中央アメリカの西海岸地域で低高度で進化したため、熱帯に適応しています。そのため、時折「低地種」とも呼ばれます。西インド原産は否定されましたが、この名前はアボカド文献にしっかりと根付いています。したがって、このカテゴリのアボカドは「熱帯低地」に適応しており、高温多湿の熱帯および亜熱帯で広く栽培されています。その範囲は霜によって制限されており、霜に対する耐性はわずかです。果実は大きく、油含量が比較的低く (「純粋な」西インド種栽培品種では 2–5%)、糖含量が高いです。熟した果実の風味は温帯アボカドとは異なり、まろやかで低地熱帯国では高く評価されています。米国フロリダでの研究は、改良された西インド種の栽培品種の選択に大きく貢献しました。これらの一部およびグアテマラ種と交配されたものは、より高い果実油含量 (6–12%) を持っています (本書第 8 章の Crane らを参照)。しかし、ほとんどの低地熱帯アボカド生産は、通常、低レベルの技術で管理される小規模農家の多作物システムで行われる非接ぎ木の苗木から得られます。この果実の非常に少量が国際貿易に入ります。
これに対して、世界貿易の観点から商業的に重要なグアテマラ種の遺伝資源は、おおむね高地の山岳雲霧林で進化しました。メキシコ種は、熱帯高地および境界的な「冷温帯」に適応しています。両種のいくつかの霜耐性は生存のための前提条件であり、特にメキシコ種にとっては重要でした。山岳斜面の良好な空気排水と常緑林の環境は、野生のアボカドの木がメソアメリカの海抜 2500〜3000 メートルの高度で生存できる理由を説明しています。「亜熱帯」アボカドが、夏雨地域の冬季または地中海気候の夏季に季節的な干ばつに耐える驚くべき耐性は、ネイティブの山岳地域での乾燥した冬と春に応答して進化したと考えられます (本書第 5 章の Wolstenholme を参照)。
グアテマラおよびメキシコ種栽培品種およびその雑種の果実は、油含量が 9〜30% と高く、糖含量が低く、「ナッツのような」風味が豊かであり、温帯アボカドよりも習得された味とされています。実際、アボカドは西洋世界で受け入れられるのが遅かったのです。事実、先住アメリカ人によるアボカドの
一般的な催淫剤特性に間接的に言及する巧妙な「逆心理」広告の後、アボカドの販売がヨーロッパで急上昇したと言われています。今日、この種のアボカドは、メキシコのミチョアカン州やドミニカ共和国、グアテマラ、およびその他の中央アメリカの高地、高地に潜在的なケニア高地のような熱帯高地で栽培されています。しかし、アボカドは南アフリカおよび東オーストラリアの一部で温暖湿潤の温帯で同様にうまく育ち、霜耐性が必要な他の大陸の高地 (例: ブラジル、アルゼンチン、および多くのアジアおよびアフリカの国々) でも潜在的に育ちます。カリフォルニア州は、海岸および沿岸の半乾燥気候で商業アボカド栽培を先駆け、イスラエル、スペイン、チリ、ペルーでのその後の農園の道を開きました。これらの産業は、今日、「ハス」と少量の「ハス」に似た新しい選択品種、および少数の「グリーンスキン」栽培品種によって支配されています。冷涼な半乾燥地域での干ばつおよび一般的なストレス耐性は、平均収量に一定のコストがかかります。これは、塩分ストレスによって悪化し、「ハス」果実が受け入れ可能な成熟度に達するためには 2 シーズン目に入る必要があることに起因します。
アボカドの木の適応戦略
「亜熱帯」アボカドのメソアメリカ山岳雨林起源は、ある程度は持続する適応戦略をもたらしました。多くのこれらが現代の農園環境では逆効果であると仮定されています (Wolstenholme と Whiley, 1999)。それらは次のように要約されます:
栄養適応戦略
– 樹木の建築構造 (Rauh 建築モデル, Halle et al., 1978) は、クライマックス森林の樹木との競争に適応しています。しかし、強い再発生の可能性があり、剪定に非常に反応します。
– 2-4 回のフラッシュで非常に活発な末端成長の可能性があり、継続的に樹木の大きさを増加させます。
– 外側の樹冠に大きく、濃緑色の二色の (グアテマラ種、メキシコ種) から淡緑色の (西インド種) 葉があります。
– 葉は比較的日陰に耐えますが、過度に日陰になると短命であり、塩分や環境ストレスにさらされると寿命が短くなります。
– 自己マルチングの木は、落葉の蓄積によって自己マルチングします。
– 酸素要件の高い浅く、効率の悪い繊維状の根が主です。
– 健康な木は、しばしば希少な栄養資源を効率的に捕捉、保持、および再利用します。
アボカドの木の生殖戦略
– 周辺の新芽の高い光要件。
– 一部の季節に周辺で非常に重い開花が見られ、寒冷および干ばつストレスによってさまざまな程度に同期します。
– 原始的な「無駄な」開花は、重要な時期に水および鉱物資源に大きな要求を課します。
– 光が制限された森林環境では、マストまたは不規則な結実が標準です。
– 小さく、個別には十分でない花は、小さな昆虫による受粉に適応しています (ネイティブ環境ではミツバチではありません)。
– 同期する二型雌雄異熟性は、義務的な異交を促進しますが、自己受粉のためのフェイルセーフを提供します。
– 花、果実、および果実の大量落下は、樹木資源および環境条件に応じて生態学的に「適応」した果実 (作物負荷) に調整されます。初期および最終的な果実の設定率は非常に低いです。
– 作物負荷は開花の重さと相関します。
– エネルギー消費 (油貯蔵) ですが、比較的鉱物消費の少ない果実。
– 集中した食糧貯蔵を持つ大きな化学的に保護された種子。
– 果実の成熟 (軟化) は木から物理的に分離された後にのみ行われます。絶滅した大型草食動物の分散エージェントへの適応。
近代的な農園の要求
上記の多くの適応が現代の農園では逆効果であり、アボカドの木が比較的新しい作物であり、まだ半家畜化されているため、平均収量がまだ比較的低いのは驚くことではありません。これはまた、油貯蔵果実のエネルギー集約的な性質と、大きく栄養豊富な種子によっても引き起こされます (Wolstenholme, 1986, 1987)。豊作 (例えば、5〜8年ごとの良好な作物) は農園の特徴ではありませんが、交互または不規則な結実は確かにあります。結果として、国家平均収量は 10 t ha−1 未満であり、良好な生産者の収量は 10〜20 t ha−1 の範囲です。低ストレス (メシック) な気候と良好な排水土壌の選択が、生産コストを削減し、平均収量を増やすための最初のステップです (本書第 5 章の Wolstenholme を参照)。ただし、経済的な考慮事項により、時には最適以下の高ストレス環境が正当化されることがあります。例えば、季節外れの果実のための高い価格などです。
農園では、アボカドの木はメシック環境で大きなサイズに成長する可能性があり、これが間隔と樹冠管理に影響を与えます。農園では森林に比べて光関係が大幅に改善され、年間の開花と収穫が可能になります。しかし、農園が成熟するにつれて、交互結実は少なくともある程度発生します。これは、樹木の生態学、発芽部位の数 (成長フラッシュ中の芽の破壊の範囲)、開花の重さ、作物負荷、農園管理、および環境の乱れに関連しています。これらのトピックは、本書の Reproductive Biology (Salazar-García et al., 第 6 章)、Ecophysiology (Schaffer et al., 第 7 章)、Irrigation and Fertilization (Lahav et al., 第 11 章)、および Crop Management (Whiley et al., 第 12 章) で徹底的に探求されています。
作物管理の観点から、樹木の間隔、トレーニング、および剪定に関するトピックは、現在、生産者およびアドバイザーの間で最も関心を引いています。新しい植栽の現在の樹木密度は、約 200 から 1666 本/ha まで変化し、一部の試験植栽では 2500 本/ha (2 × 2 m 間隔) です。この変動は、異なる栽培地域での樹木の活力の違いや、環境条件、熟練した労働力、資本、研究、および企業家精神によって正当化される管理の集約度の違いを反映しています。温暖湿潤の温帯気候と良好な土壌は、非常に高密度の植栽を除外
します。半乾燥および乾燥気候を含む気候は、木の活力を低下させ、樹木の間隔を縮小することを正当化します。それでも、段階的な樹木間引きなしで高い樹木密度を維持するには、樹木のトレーニング、定期的な剪定、およびバイオレギュラントの使用を含む非常に集中的な管理が必要です。これらの進化する概念を利用した超高密度農園の先駆的な作業が現在、チリで行われています (Gardiazabal et al., 2011; Mena et al., 2011)。
熱帯および亜熱帯の緯度全体で、人口増加と地球規模の気候変動は、大気中の CO2 濃度の上昇、気温の上昇、低品質の水、干ばつ、洪水、および塩水侵入のような破壊的な環境現象の増加をもたらすと予測されています (Vörösmarty et al., 2000; IPPC 2007; Koning et al., 2008; IPPC, 2009)。疑いなく、アボカドの生産性はこれらの変化する環境シナリオによって影響を受けるでしょう。制御環境研究では、一定温度下での大気中 CO2 濃度の増加がアボカドの光合成と成長を増加させることが示されました (Schaffer et al., 1999)。しかし、アボカドの生産性に対するより大きな影響は、気温の変化によって成長と発達のいくつかの側面に影響を与えることです。例えば、花の形成や熱ストレスです。したがって、環境要因がアボカドの生物学に与える影響を理解することは、変化する気候の下で最大の生産性を達成するために木や農園を操作するためのフレームワークを提供します。また、干ばつ、洪水、および塩分の増加などの環境リスクの影響を軽減するために、改良された品種と台木の開発や、より効率的な農園管理が必要になるでしょう。したがって、今日のアボカド生産者は大きな課題に直面していますが、長期的には継続的な研究によってのみ解決されるでしょう。本書のいくつかの章 (例えば、Wolstenholme ら, 第 5 章; Salazar-García ら, 第 6 章; Schaffer ら, 第 7 章) では、基本的なアボカド生物学に対する気候変数の影響について詳細な議論が含まれています。一部の章 (例えば、Lahav ら, 第 11 章; Whiley ら, 第 12 章) では、環境変数に関連した木および農園管理に関する実用的な議論も提供しています。主な技術的課題は、最終的には、気候変動に対応するための新しいエリート品種と台木の育種によって解決されるでしょう (本書第 4 章の Lahav と Lavi を参照)。
急速に発展している新しい基礎研究技術の議論は、本書の分類学と植物学に関する章 (Chanderbali ら, 第 3 章)、遺伝学と育種 (Lahav と Lavi, 第 4 章)、およびバイオテクノロジー (Pliego-Alfaro ら, 第 10 章) に含まれています。いったん解決困難だった問題に関しても顕著な進展が見られ、必要な人材と施設への十分な投資があれば、驚異的な進歩が可能です。この基礎研究は、業界が競争に追いつくだけでなく、競争優位を獲得するための手助けとなります。
主要な病害と害虫
どの果樹作物も、多かれ少なかれ病害虫のスイートに悩まされます。最も破壊的な病害は間違いなく Phytophthora 根腐れ病であり、ほぼ世界中に分布しています。アボカドの進化の過程でこの病原菌が自然に存在しなかったため、アボカドは Phytophthora 根腐れ病に対する完全な耐性を進化させませんでした。この病原菌は、排水の悪い農園を壊滅させる可能性があり、温暖湿潤地域では特に破壊的です。効果的な化学的制御は、1970 年代半ば以降ほとんどの国で利用可能になりましたが、長期的には、統合管理プログラムを伴わない限り不十分です (Pegg, 2010)。いくつかのクローン台木には部分的な耐性がありますが、これだけでは不十分です。過去 10 年間で、この病害の管理に関する重要な進展が見られましたが、本書第 13 章の Dann らによって他の病害についても議論されています。
アボカド農園は、印象的な害虫やダニの害にさらされる可能性があります。しかし、これらのほとんどは通常、軽微な関心事または散発的なものであり、通常、化学的な是正措置を必要としません。どの国でも主要な害虫は注意深い統合害虫管理プログラムで制御する必要があります。これらの一部は、潜在的な輸入国における植物検疫障壁のために重要となります。例えば、果実バエや偽幼虫などです。本書第 14 章の Peña らは、特定の国や地域に特有の多くの害虫やダニ害について議論しています。
収穫前の栽培活動は、成功した生産者になるための要件の一部にすぎません。効率的な収穫、梱包、輸送、および加工は、アボカド生産において主要な技術的課題を呈します。ポストハーベスト研究は、特に南北半球間の長距離輸出において、アボカド生産の極めて重要な要素となっています。限られたポストハーベスト寿命と、ポストハーベスト病害および障害に対する感受性により、アボカドは最も輸出が難しい果実の一つです。裕福な買い手は品質と高い基準を要求し、専用の研究努力にもかかわらず、現在の栽培品種では内部果実の品質を常に保証することはできません。現在の最良のポストハーベストプラクティスは、本書第 15 章の Hofman らによって議論されています。
アボカドマーケティングに関する章を含める予定でしたが、この急速に変化する分野は、現在の経済状況に迅速に反映および対応できる他のメディアで最も適切に扱われるでしょう。それにもかかわらず、マーケティングおよび経済的考慮事項が最終的な決定を下すことが常に真実です。園芸的に望ましいことは、経済的に正当化されないかもしれません。持続可能な農業企業の定義には経済が含まれるべきであり、生産者が利益を上げることが必須です。収入が支出を上回る必要があります。市場調査は現在および将来のために不可欠です。幸いなことに、管理の行き届いた現代の商業果樹園は、環境影響、食品安全、および農業持続可能性の他の指標に関して多くの利点があります。特に、低農薬使用、害虫および病害の統合管理の強調、マルチの広範な使用、非常に栄養価の高い製品の安全記録、小規模な企業の多さ、および農園の集
中的な管理などです。アボカド農園は、真に持続可能な企業を開発するためのモデルとなり得ます。しかし、一部の地域では、投入コストが卸売果実価格の上昇速度を上回る傾向があり、アボカド生産の長期的な持続可能性に関する懸念が生じています。
結論
最新のゲノム技術を使用した分類学研究は、アボカドが被子植物の起源に近い「基部系統 (マグノリアクレード)」を代表する「進化の先端」であることを確認しました (Chanderbali ら, 2008)。実際、それは遺伝的な花の進化と基部被子植物の果実の発達の基本的な研究を引きつける確立されたモデル果樹作物となっています。
アボカドは少なくとも 9000 年間、人間によって評価されてきました。現代の優れた栽培品種の選択および栄養繁殖は、過去 110 年間、フロリダの「ポロック」と「トラップ」、およびカリフォルニアの「フエルテ」に始まりました。世界的な作物として、アボカドは重要で急速に拡大している果樹作物ですが、年間約 360 万トンの生産量では、ブドウ、柑橘類、バナナ、リンゴなどと比較するとまだ第二段階の果実です。メキシコは最大の生産国であり、次いでチリ、米国、およびペルーが続きます。世界の作物の約 20% が国際貿易に回されており、主要な輸入国は米国および EU です。過去 10 年間の生産の急速な拡大は、段階的な植物検疫規制の緩和によって米国市場の開放に起因しています。
アボカドはメソアメリカに原産であり、今日「熱帯」低地タイプ (西インド種) は本物の熱帯国および温暖な亜熱帯地域で栽培されています。「亜熱帯」タイプは、高地山岳雲霧林のグアテマラ種および/またはメキシコ種に基づいており、今日、世界の温暖から冷温帯および地中海気候で広く栽培されています。「ハス」品種が支配的です。野生のアボカドの木の明らかな栄養および生殖戦略は、砂糖貯蔵果樹作物と比較して低い平均収量を説明するのに役立ちます。
主要な技術的課題は、温暖湿潤地域での生産者の平均収量が 8〜12 t ha−1 であり、30 t ha−1 を超える潜在的な収量があることです。現在の栽培品種、台木、および技術では、交互または不規則な結実が依然として課題です。Phytophthora 根腐れ病は、アボカド農園の主要な病害であり、統合管理プロトコルを必要とします。主な亜熱帯品種「ハス」は、新しい「ハス」に似た選択品種によって補完されていますが、亜熱帯および低地熱帯の両方に改良されたエリート品種が必要です。Phytophthora 根腐れ病に耐性のあるクローン台木で進展が見られますが、真に耐性のあるエリート台木が必要です。アボカド農園の最良の管理プラクティスはさまざまであり、樹木の間隔、トレーニングおよび剪定、交互結実の制御に関して進化を続けています。さらに、品種育成、栽培品種選択、および農園管理は、気候変動に適応する必要があります。
続く 14 章は、9 つのアボカド生産国の 45 人の著名なアボカド研究者の研究、観察、および解釈に基づいて、アボカド科学の最新 (2012 年初頭) の世界的な状況を示しています。
参考文献
– Barlow, C. (2000). The Ghosts of Evolution. Basic Books, New York, 291 pp.
– Chanderbali, A.S., Albert, V.A., Ashworth, V.E.T.M., Clegg, M.T., Litz R.E., Soltis, D.E., Soltis, P.S. (2008). Persea americana (avocado): bringing ancient flowers to fruit in the genomics era. BioEssays 304, 386–396.
– Chanderbali, A.S., Albert, V.A., Leebens-Mackd, J., Altmane N.S., Soltis, D.E., Soltis, P.S. (2009). Transcriptional signatures of ancient floral developmental genetics in avocado (Persea americana; Lauraceae). Proceedings of the National Academy of Sciences 106, 8929–8934.
– Fairchild, D. (1945). Personal recollections of George B. Cellon, horticultural pioneer of south Florida. Proceedings of the Florida State Horticultural Society 58, 205–209.
– Gama-Campillo, L., Gomez-Pompa, A. (1992). An ethnoecological approach for the study of Persea: a case study in the Maya area. Proceedings of the Second World Avocado Congress, University of California, Orange, California, pp. 11–17.
– Gardiazabal, F., Mena, F., Magdahl, C., Adriazola, C., Torres, J. (2011). Neuvos sistemas de poda en arbo-les adultos de paltos (Persea americana Mill.), cv Hass. VII World Avocado Congress, Cairns, Australia. Available at: [http://worldavocadocongress2011.com/userfiles/file/Francisco%20Gardiazabal%201500-1520.pdf](http://worldavocadocongress2011.com/userfiles/file/Francisco%20Gardiazabal%201500-1520.pdf) (accessed 7 May 2012).
– Hallé, F., Oldeman, R.A.A., Tomlinson, P.B. (1978). Tropical Trees and Forests: An Architectural Analysis. Springer, Berlin, 441 pp.
– Heywood, V.H. (ed.) (1978). Flowering Plants of the World. Oxford University Press, Oxford, 336 pp.
– Imbert, E. (2010). The world avocado market in 2009–10: growth all round. FruiTrop no. 181, 17–25.
– IPPC (Intergovernmental Panel on Climate Change) (2007). Climate Change 2007: The Physical Science Basis. Cambridge University Press, Cambridge.